
{kind=link}
棉花幼苗钾吸收的系统反馈调节初步研究
引用本文
王晔, 田晓莉. 棉花幼苗钾吸收的系统反馈调节初步研究. 作物学报, 2013, 39(10): 1843-1848
[WANG Ye, TIAN Xiao-Li. Systemic Feedback Regulation of K+ Uptake in Cotton at Seedling Stage . Acta Agronomica Sinica, 2013, 39(10): 1843-1848]
Permissions
[WANG Ye, TIAN Xiao-Li. Systemic Feedback Regulation of K+ Uptake in Cotton at Seedling Stage . Acta Agronomica Sinica, 2013, 39(10): 1843-1848]
Copyright©2013, Editorial office of Acta Agronomica Sinica
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
棉花幼苗钾吸收的系统反馈调节初步研究
摘要
以中棉所41和辽棉17为材料, 采用单接穗双砧木嫁接的方法构建分根体系, 在营养液培养条件下研究棉花幼苗钾吸收的系统反馈调节。分根处理6 d后, 高钾侧(2.50 mmol L-1)根系(Sp.+K)的吸收能力受到低钾侧(0.01 mmol L-1) K+需求信号的诱导, 吸收动力学参数
关键词:
棉花; 分根; 钾吸收; 反馈调节
Systemic Feedback Regulation of K+ Uptake in Cotton at Seedling Stage
Abstract
This hydroponic split-root experiment was conducted to elucidate the long-distance systemic regulation of K+ uptake in cotton (
Keyword:
Cotton; Split-root; K uptake; Systemic regulation
引言
土壤中矿质养分的有效性处于波动状态, 植物不仅以高效的感应系统(局部信号)感知外部养分浓度的变化, 还以内部系统信号报告各器官的养分状态, 从而精细调节养分的运输和代谢, 使根系的吸收能力与植株需求相匹配[1,2]。系统信号包括根-冠和冠-根两种长距离信号, 分别传递根系感受到的养分变化信息和地上部的养分需求信息, 冠-根信号又称反馈信号。近20余年来, 养分吸收的局部调节研究在细胞水平和分子水平上取得较快进展[3,4,5], 但对长距离系统调节仍所知甚少[6,7]。
K+是植物必需的大量营养元素之一, 占植物干重的2%~10%[8], 在酶的活化、膜运输、阴离子中和及渗透调节等方面发挥重要作用[9], 并显著影响光合作用和同化产物运输等生理过程, 最终导致植物生长和发育的变化[10]。棉花属于喜钾作物[11], 但其根系在耕层和表层土壤的分布比较稀疏[12,13], 因而对缺钾比较敏感[14]。K+供应不足时棉花发生一系列生理异常[15], 产量和品质也受到较大影响[16]。
关于植物K+吸收的研究起步较早, 在20世纪90年代之前形成了两种相反的意见。一种认为根系组织的K+浓度([K+])对K+吸收起主要调节作用[17], 另一种认为地上部是影响K+吸收的主要因素[18,19]。20世纪90年代后, K+吸收研究集中在K+通道和运输载体的鉴定及细胞水平调控网络的揭示上[5], 仅有个别报道涉及长距离调控[20]。本文采用分根和去根等方法研究了棉花K+吸收的长距离系统反馈调节现象, 一方面为明确K+吸收的系统调节提供试验证据, 另一方面为制定适宜栽培措施缓解棉花的缺钾问题提供理论依据。
1 材料与方法
1.1 试验材料及处理
供试品种中棉所41和辽棉17种子分别由中国农业科学院棉花研究所和辽宁省农业科学院经济作物研究所提供。试验于中国农业大学光照培养室进行, 光照强度为600 μmol cm-2 s-1, 光照/黑暗时间为12 h/12 h, 昼/夜温度为(30±2)℃/(22±2)℃, 相对湿度为70%~80%。种子经9%双氧水消毒15~20 min后用流水清洗数遍, 在去离子水中浸种12 h, 露白后播种于不含K+的沙床, 出苗2 d后转移至K+浓度为2.50 mmol L-1的1/2改良Hoagland营养液中[21]。
采用“单接穗双砧木”自身嫁接法[21]构建分根植株(图1), 嫁接位点位于下胚轴。砧木和接穗在上述营养液中培养5 d, 至第1片真叶出现时嫁接, 嫁接苗在高湿(相对湿度75%左右)、弱光(80 μmol cm-2 s-1)环境下缓苗7 d, 后移至600 μmol cm-2 s-1光强下培养。
1.2 试验设计
选择生长一致的嫁接棉株, 将2个砧木分别放入相邻的2个用锡箔纸包裹的300 mL玻璃管中(图1), 在2.50 mmol L-1 K+营养液中培养至三叶期开始分根处理。高钾侧(Split-root +K, 简写为Sp.+K)K+浓度为2.50 mmol L-1, 低钾侧(Split-root -K, 简写为Sp.-K)K+浓度为0.01 mmol L-1, 并分别设高钾对照(Control +K, 简写为C.+K, 两侧K+浓度均为2.5 mmol L-1)和缺钾对照(Control -K, 简写为C.-K, 两侧K+浓度均为0.01 mmol L-1), 处理6 d后取样。
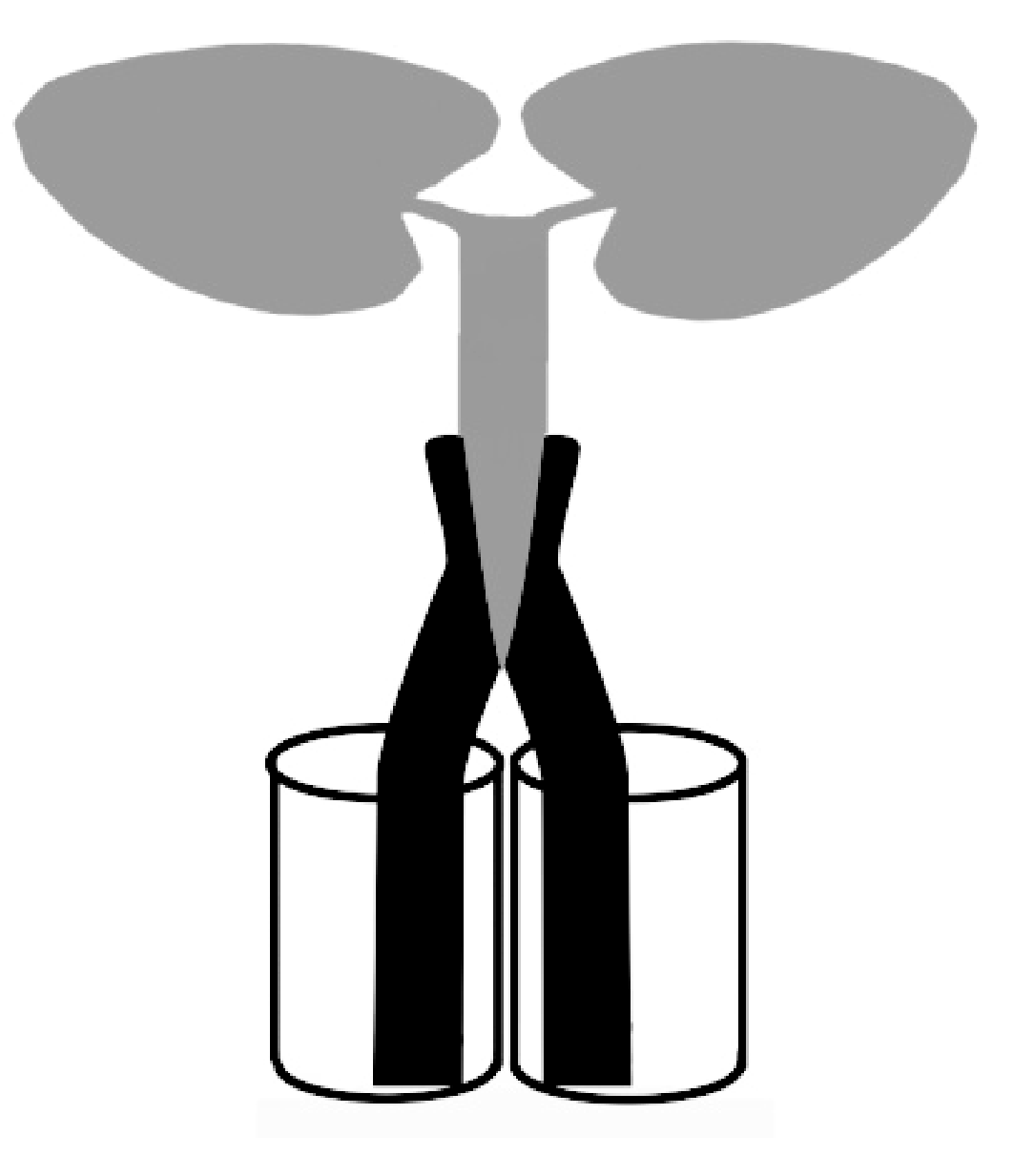 | 图1 单接穗双砧木分根系统Fig. 1 Split-root plant of one scion/two rootstocks |
此外, 中棉所41在分根处理的基础上开展去根试验, 即在分根处理3 d后剪除Sp.-K侧根系, Sp.+K侧则正常供K+, 继续培养3 d后取样, 该处理表示为Remove the split-root-K, 简写为Re.Sp.-K。每处理重复3次, 每天更换一次营养液, 24 h通气。
1.3 取样及测定
1.3.1 干重和钾含量 用去离子水反复冲洗根系, 然后将植株在嫁接位点处分为接穗和2个砧木共三部分, 置105℃鼓风干燥箱杀青15 min, 在80℃下烘至恒重后记录重量。将烘干样品研成粉末, 用1 mol L-1 HCl振荡浸提8 h, 采用原子吸收分光光度计(SpectAA-50/55, Varian, Australia)测定滤液的钾 浓度。
1.3.2 K+吸收动力学参数测定 将各处理植株先在含0.2 mmol L-1 CaSO4、pH为5.7的饥饿液中饥饿24 h(每12 h换一次饥饿液), 然后用去离子水冲洗根系, 将2个砧木分别移入用锡箔纸包裹并盛有250 mL耗竭液的2个相邻玻璃瓶中(图1), 每瓶2株, 重复3次。耗竭液起始K+浓度为0.25 mmol L-1, 分别于5、15、25、40、70、100、130、160、190、250、310、370、490、610、670、730、850、970、1150、1330、1690和1930 min取耗竭液, 每次吸取1 mL, 同时补充1 mL原始耗竭液。测定各次所取耗竭液的K+浓度, 绘制K+耗竭曲线。根据蒋廷惠等[22]的方法计算K+最大吸收速率( Imax)、根系对离子的亲和常数( Km)和吸收速率为零时的外界钾离子浓度( Cmin)。
1.4 数据统计
各试验均独立重复3次以上, 各次结果趋势一致, 文中所用数据为其中具有代表性的一次。用SAS统计软件(V8, SAS Institute Inc., Cary, USA)的ANOVA法进行差异显著性分析( P=0.05), 用Duncan’s法进行多重比较。
2 结果与分析
2.1 干物重和钾含量
表1结果显示, 两品种C. -K的生长及钾含量显著低于C.+K, 其中中棉所41地上部干重和钾含量分别降低40%和14%, 根系干重和钾含量分别降低24%和20%; 辽棉17地上部干重和钾含量分别降低33%和8.5%, 根系干重和钾含量分别降低21%和18%。两品种Sp.+K-K的地上部干重和钾含量显著高于C.-K(辽棉17钾含量除外), 与C.+K相比有降低趋势但差异不显著(中棉所41钾含量除外), 说明局部根系供K+在短时间内(6 d)基本可满足棉株正常生长的需要。两品种Sp.+K侧的根系干重和钾含量低于C.+K, Sp.-K侧的根系干重和钾含量高于C.-K, Sp.+K侧有高于Sp.-K侧的趋势, 但这些差异均不 显著。
中棉所41分根处理3 d后去除Sp.-K侧根系(Re.Sp.-K), 继续培养3 d, 发现地上部干重和钾含量较Sp.+K-K有所增加, 但差异未达显著水平; 保留的高钾侧根系[Sp.+K (Re.Sp.-K)]的干重和钾含量也较Sp.+K增加, 其中干重差异达到显著水平, 与C.+K基本相当。
| 表1 棉花分根幼苗(两侧均等或非均等供K+)地上部和根系的干重及钾含量 Table 1 Dry weight and K+ concentration of both shoot and root of cotton split-root seedlings |
| 表2 棉花分根幼苗(两侧均等或非均等供K+)的K+吸收动力学参数 Table 2 Kinetic parameter of K+-uptake in cotton split-root seedlings |
2.2 根系K+吸收动力学参数
从表2可以看出, 低钾对照(C.-K)的根系吸K+能力显著高于高钾对照(C.+K)。中棉所41和辽棉17 C.-K的 Imax较C.+K分别高2.9倍和2.4倍, Km较后者分别低51%和33%, Cmin仅相当于后者的8%和26%。分根处理6 d后, 中棉所41和辽棉17 Sp.+K侧根系的 Imax与C.+K相比分别显著提高92%和79%, Sp. -K侧根系的 Imax与C.-K相比则分别降低26%和40%; 两品种Sp.-K侧的 Imax均高于Sp.+K侧, 但差异不显著。Sp.+K侧根系的 Km和 Cmin与C.+K相比显著降低了17%~29%和38%~49%, Sp.-K侧根系的 Km和 Cmin与C.-K相比显著增加了34%~41%和216%~531%; 两侧的 Km和 Cmin无显著差异。
中棉所41分根处理3 d后去除Sp.-K侧根系, 保留的Sp.+K侧根系[Sp.+K(Re.Sp.-K)]的吸K+能力在继续培养3 d后较Sp.+K (不去除Sp.-K侧根系)下降, 基本恢复到C.+K的水平, 其中 Imax虽然仍较C.+K高50%但差异不显著, Km和 Cmin则与C.+K相当。
3 讨论
绝大多数研究证明, 矿质养分吸收的长距离反馈调节现象普遍存在。拟南芥分根植株一侧根系处于NO3-饥饿状态时, 另一侧根系(供应NO3-)的NO3-内流速率和At NRT2.1 (编码高亲和NO3-运输载体)转录丰度上调[23]。将分根试验与转录谱检测相结合, 发现蒺藜苜蓿NO3-和NH4+的吸收及根瘤N2的固定均受到系统反馈调节, 且每一种N源都有其特异的系统调控网络[24]。植株对Pi的需求通过供Pi部分根系得到满足后, 缺Pi部分根系Pi转运蛋白的表达不再受低Pi诱导[25,26,27]。植物的部分根系处于缺Fe条件下时, 可诱导另一部分供Fe根系的Fe(III)还原酶活性和/或质子分泌增强[28,29,30]。
Claassen和Barber[18]及Drew和Saker[19]的早期分根试验证明, K+吸收能力也受到地上部的反馈调节。分根处理10 d, 玉米幼苗供K+部分根系的K+内流速率较整根供K+增加2.6倍[18]; 大麦分根处理3~4 d, 供K+部分根系42K和86Rb的吸收速率较整根供K+提高近一倍[19]。但近期的胡椒分根试验表明, 处理3 d后缺K+侧根系的高亲和K+吸收与供K+侧的K+状况无关, 提示地上部调节高亲和K+吸收的可能性不大[20]。本试验棉花幼苗分根处理6 d, 高K+侧根系(Sp.+K)的 Imax在低K+侧(Sp.-K)发出的缺K+信号诱导下较高钾对照(C.+K)增加79%~92%; Sp.-K侧根系的 Imax在高K+侧(Sp.+K)发出的供K+信号作用下较低钾对照(C.-K)降低26%~40%; 分根处理3 d后去除缺钾感应器官Sp.-K侧根系, 再经过3 d后Sp.+K侧根系的 Imax虽然仍高于C.+K但差异不显著, Km和 Cmin则恢复到C.+K的水平。这些结果为棉花幼苗K+吸收的反馈调节提供了可靠的证据, 支持早期的研究结果[18,19]。
最近, Ruffel等[31]的拟南芥分根试验显示, 植物至少存在两种遗传上独立的系统信号, 分别报告植株的N供应和N需求状况。分根处理供N侧的侧根长度大于高N对照, 他们认为这是长距离N-需求信号(由缺N侧发出)和局部NO3-供应共同作用的结果; 分根处理缺N侧的侧根长度小于缺N对照, 则可能与供N侧发出的N-供应信号(抑制信号)有关。本试验结果表明, 分根处理Sp.+K根系的 Imax较C.+K增加了79%~92%, 而Sp.-K根系的 Imax较C.-K仅下降27%~40%。我们推测, Sp.-K侧根系发出的K+-需求信号较强, 因而Sp.+K侧根系的吸收速率得到较大提升; 来自Sp.+K侧的K+-供应信号较弱, 所以Sp.-K侧根系的 Imax变化较小。与 Imax相反, 分根处理Sp.+K侧 Km和 Cmin与C.+K的差异小于Sp.-K侧根系与C.-K的差异(Sp.+K侧的 Km和 Cmin值较C.+K分别下降17%~29%和38%~49%, 而Sp.-K侧根系的 Km和 Cmin值较C.-K分别增加34%~41%和216%~ 528%), 提示棉花根系K+通道/运输载体的数量与亲和性可能受不同系统信号的调节。
由于不同物种根系中的[K+]与K+内流速率呈相反关系, 早期研究认为根系细胞[K+](可能是细胞质中的[K+])对K+内流进行负反馈变构调节[17,32,33,34,35]。Claassen和Barber[18]的结果则相反, 他们将分根试验与K+浓度梯度试验、K+饥饿不同时间试验的数据综合起来计算, 发现地上部[K+]较根系[K+]与K+最大吸收速率( Imax)的关系更为密切, R2达到0.74。然而, 此后细胞水平和分子水平上的研究既未明确根系细胞中的K+在调控K+通道/运输载体表达中有何作用[5], 也未进一步揭示地上部K+调控根系吸收能力的生理机制。本试验中棉所41分根处理Sp.+K侧的 Imax较C.+K提高92%, 但该侧根系和地上部的[K+]较后者仅分别降低10%和6%; 辽棉17分根处理Sp.+K侧的 Imax较C.+K提高79%, 但该侧根系和地上部的[K+]与后者基本相当。这些结果表明, 根系和地上部的K+未直接参与K+吸收的调控。
与局部供N和供Pi不同, 根系在局部供K+条件下不发生明显的补偿生长[36,37,38]。与此一致, 本试验分根处理Sp.+K侧根系的干重低于或显著低于C.+K两侧的平均值, 与Sp.-K侧根系基本相当, 未表现出补偿生长现象。De Jager[39]和Ma等[40]的结果则相反, 他们发现局部供K+对根系生长的刺激效应分别与局部供NO3-和Pi相当, 但前提是缺K+部分完全不供K+或长时间分根处理后地上部的[K+]明显降至最适状态以下。本试验分根处理Sp.-K侧的供K+浓度为0.01 mmol L-1, 处理结束时(持续6 d)地上部的[K+]与C.-K均处于较高水平(30 mg g-1左右)。如果延长棉花幼苗分根处理时间, 或将Sp.-K侧的供K+浓度降为0, Sp.+K侧根系是否会出现补偿生长则未可知。
4 结论
分根处理后高钾侧(2.50 mmol L-1)根系受到低钾侧(0.01 mmol L-1) K+需求信号的诱导其 Imax较高钾对照增加, 低钾侧根系则受到高钾侧K+供应信号的抑制其 Imax较低钾对照降低; 去除低钾侧根系, 保留的高钾侧根系因失去K+需求信号的诱导其 Imax较完整分根植株有所下降。根系和地上部组织的[K+]与K+吸收的这种系统反馈调节没有直接关系。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|